
Review ArticleOpen Access, Volume 3 Issue 2
Cytokine and chemokine modulation for immune reprogramming in cancer and neurodegeneration: A comprehensive therapeutic review
Neha Anees1; Hammad Ali Hassan2; Ayesha Siddiqa1; Umer Anayyat3; Aliza Fatima4; Yang Yi5#; Xiaomei Wang3#*
1Department of Zoology, Wildlife and Fisheries, University of Agriculture, Faisalabad, Pakistan.
2Department of Neurosurgery, University of Alabama at Birmingham, Birmingham, AL, USA.
3School of Biomedical Engineering Sciences, Shenzhen University, Shenzhen, Guangdong, 518061, People’s Republic of China.
4Department of Biochemistry, The University of Faisalabad, Faisalabad, Pakistan.
5Department of Biochemistry and Molecular Biology, Shenzhen University Medical School, China
#These authors have been equally contributed to this article.
*Corresponding author: Xiaomei Wang
School of Biomedical Engineering Sciences, Shenzhen University, Shenzhen, Guangdong, 518061, People’s Republic of China.
Email: xmwang@szu.edu.cn
Received : Nov 10, 2025 Accepted : Dec 17, 2025 Published : Dec 23, 2025
Epidemiology & Public Health - www.jpublichealth.org
Copyright: Wang X © All rights are reserved
Citation: Anees N, Hassan HA, Siddiqa A, Anayyat U, Fatima A, et al. Cytokine and chemokine modulation for immune reprogramming in cancer and neurodegeneration: A comprehensive therapeutic review. Epidemiol Public Health. 2025; 3(2): 1083.
Abstract
Immune reprogramming, the dynamic process by which immune cell phenotypes and functions adapt in response to microenvironmental signals, plays a central role in both cancer progression and neurodegenerative disease pathogenesis. Cytokines and chemokines serve as critical orchestrators of this process, functioning as signaling molecules that coordinate immune system responses. In cancer, malignant cells commandeer these pathways to establish an immunosuppressive Tumor Microenvironment (TME) that recruits pro-tumorigenic cells while inhibiting anti-tumor immunity. Conversely, in neurodegenerative disorders, aberrant cytokine and chemokine signaling drives chronic neuroinflammation, ultimately resulting in progressive neuronal damage.
This comprehensive review synthesizes current literature on immune reprogramming mediated by cytokines and chemokines, with emphasis on therapeutic applications in both disease contexts. We examine diverse therapeutic modalities for modulating these pathways, ranging from traditional recombinant cytokines to contemporary approaches including monoclonal antibodies and novel delivery systems. Critical challenges impeding clinical translation are discussed, including systemic toxicity, context-dependent functions of these molecules, and the formidable barrier posed by the blood-brain barrier in neurological applications.
Finally, we outline future research priorities, emphasizing the need for highly specific targeted therapies and rationally designed combination strategies to restore appropriate immune balance in combating these devastating diseases.
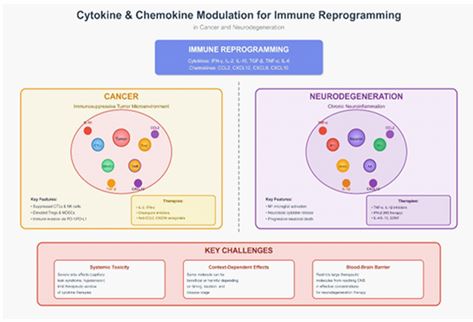
Introduction
The immune system comprises a complex network of cells, tissues, and organs that collectively defend the body against diverse threats, including pathogenic microorganisms and ma lignant cells. Rather than functioning as a static defense mecha nism, the immune system exhibits remarkable plasticity through a process termed immune reprogramming [45]. This process refers to dynamic alterations in immune cell phenotype and function in response to microenvironmental cues, representing an essential adaptive mechanism that becomes detrimental in diseases such as cancer and neurodegeneration [44].
In cancer, immune reprogramming serves as a critical mech anism of immune evasion. Malignant cells possess the capac ity to reshape the Tumor Microenvironment (TME), converting immune responses from anti-tumorigenic to pro-tumorigenic. This transformation involves suppressing Cytotoxic T Lympho cytes (CTLs) and Natural Killer (NK) cells while simultaneously recruiting and activating immunosuppressive populations, in cluding regulatory T cells (Tregs) and Myeloid-Derived Suppres sor Cells (MDSCs). This immune dysfunction enables tumors to evade immunosurveillance, facilitating uncontrolled prolifera tion and metastasis [46].
Neurodegenerative diseases, particularly Alzheimer’s Dis ease (AD) and Parkinson’s Disease (PD), similarly exhibit patho logical immune reprogramming that drives chronic neuroinflam mation. The Central Nervous System (CNS), once considered an immune-privileged site, is now recognized as being actively surveilled by resident immune cells, particularly microglia, the brain’s resident macrophages [49]. In neurodegenerative condi tions, microglia transition from their homeostatic surveillance state to a reactive inflammatory phenotype, releasing pro-in flammatory mediators that contribute to neuronal damage and disease progression. Understanding these complex immuno logical changes is essential for developing effective therapeutic interventions.
Cytokines and chemokines represent two critical families of signaling molecules that orchestrate immune reprogramming [51]. Cytokines are small secreted proteins that mediate and regulate immunity, inflammation, and hematopoiesis [94]. This diverse group includes Interleukins (ILs), Interferons (IFNs), and Tumor Necrosis Factors (TNFs), each performing distinct func tions in cellular communication. Chemokines constitute a spe cialized subset of cytokines that primarily function as chemoat tractants, directing immune cell migration to specific anatomical locations [43]. The intricate interplay between cytokines and chemokines guides immune cells to sites of infection, injury, or disease. Under physiological conditions, this communica tion network ensures prompt and effective immune responses [47,50]. However, in pathological states, these signaling path ways become dysregulated with profound consequences.
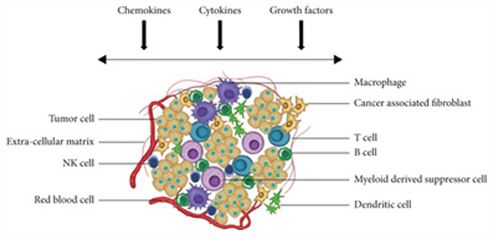
In cancer, tumors exploit cytokine and chemokine signaling to recruit pro-tumorigenic immune cells, induce angiogenesis, and suppress anti-tumor immunity, as illustrated in Figure 1. Similarly, in neurodegeneration, aberrant chemokine and cyto kine gradients can trigger chronic microglial activation and pro mote infiltration of peripheral immune cells into the CNS, in tensifying neuroinflammation and accelerating neuronal death [55].
This schematic illustrates how tumor cells and stromal com ponents secrete various signaling molecules that collectively shape the immunosuppressive TME. These factors coordinate multiple processes including recruitment of pro-tumorigenic immune cells, promotion of angiogenesis, inhibition of anti tumor immunity, and facilitation of tumor cell survival and pro liferation.
Given the central importance of cytokines and chemokines in shaping pathological immune responses in both cancer and neurodegeneration, targeting these pathways represents a promising therapeutic strategy. Over the past decade, numer ous studies and clinical trials have emerged aimed at modulat ing these signaling molecules. This review provides a compre hensive analysis of current knowledge regarding cytokine and chemokine-mediated immune reprogramming in these two distinct yet interconnected disease areas.
We first examine the specific roles of these signaling mol ecules in the tumor microenvironment and the CNS, and how they contribute to disease progression. Subsequently, we dis cuss various therapeutic strategies that have been developed, including recombinant cytokines, small molecule inhibitors, and monoclonal antibodies. We critically evaluate both the clinical successes and, more importantly, the challenges and limita tions of these treatments, including systemic toxicity and the complex context-specific actions of these molecules. Finally, we present perspectives on future research directions and how next-generation targeted therapies might be formulated to ef fectively reprogram the immune system for therapeutic benefit.
Immune reprogramming in cancer
Tumor microenvironment and immune evasion
The Tumor Microenvironment (TME) constitutes a complex ecosystem comprising tumor cells, immune cells, fibroblasts, endothelial cells, and the extracellular matrix. This intricate network serves as a major determinant of tumor progression and treatment response [102]. To evade immune detection and subsequent elimination, tumor cells have evolved sophisticated mechanisms to manipulate the TME, generating an immuno suppressive milieu that promotes tumor survival and expansion.
Immune evasion mechanisms primarily function by suppress ing anti-tumor immune responses through multiple strategies. A key mechanism involves activation of immune checkpoints, including the PD-1/PD-L1 axis, which induces T cell exhaustion and functional impairment [18,52].
Beyond immune checkpoint engagement, the TME harbors high concentrations of immunosuppressive cell populations. Myeloid-Derived Suppressor Cells (MDSCs) and Tumor-Associ ated Macrophages (TAMs) are particularly adept at promoting tumor growth through secretion of immunosuppressive cyto kines and growth factors [58,97]. These cells not only suppress effector T cell activity but also stimulate angiogenesis and facili tate tissue remodeling, further contributing to tumor progres sion (Mano et al. 2013). Additionally, regulatory T cells (Tregs) accumulate within the TME, where they actively suppress anti tumor immunity through direct cell-to-cell contact and release of inhibitory cytokines (Tommasin et al. 2017).
Cytokine and chemokine modulation in tumor immunity
Dysregulated cytokine and chemokine signaling lies at the heart of the immunosuppressive TME. Classical pro-inflamma tory cytokines such as IFN-gamma and IL-12, typically associated with robust anti-tumor immunity, are frequently suppressed in the TME [90]. Conversely, the TME exhibits elevated concentra tions of immunosuppressive cytokines, including IL-10 and TGF beta [65]. These molecules not only inhibit anti-tumor immune cell activation and proliferation but also promote differentiation of MDSCs and Tregs, further reinforcing the immunosuppres sive environment [59].
Chemokines play a crucial role in determining the cellular composition of the TME. Tumor cells and stromal cells secrete distinct chemokines that selectively recruit immunosuppressive cell types while discouraging infiltration of anti-tumor effectors, as demonstrated in Figure 1.
The diagram highlights how this complex architecture en ables tumor expansion and creates a hypoxic core. The tumor orchestrates immune evasion by recruiting immunosuppres sive cells that inhibit anti-cancer responses, thereby facilitat ing tumor survival and growth. Key features include the spatial organization of different cell types, the formation of abnormal vasculature, and the establishment of gradients that promote tumor progression while suppressing effective immune surveil lance.
For example, CCL2 has been shown to recruit macrophages into the tumor, where they undergo reprogramming into pro tumorigenic TAMs. Similarly, the CXCL12/CXCR4 chemokine axis plays a pivotal role in tumor cell proliferation, survival, and me tastasis by attracting various stromal and immune cell popula tions to the tumor site (Zu et al. 2020).
Therapeutic strategies targeting cytokine and chemokine signaling
Given their central role in shaping the TME, cytokines and chemokines represent attractive targets for cancer therapy. Therapeutic approaches can be broadly classified into two cat egories: direct modulation of cytokine or chemokine levels, and indirect intervention through receptor targeting [63]. Recombi nant cytokines, including IL-2 and IFN-alpha, were among the earliest immunotherapies applied in cancer treatment. While these agents have demonstrated clinical benefit in select malig nancies, their widespread application has been limited by sig nificant systemic toxicity and short half-lives.
More recently, targeted approaches have gained promi nence. These include monoclonal antibodies designed to neu tralize immunosuppressive cytokines such as IL-10 or TGF-beta, as well as antibodies targeting chemokines that recruit pro-tu morigenic cells [61]. Novel strategies employ oncolytic viruses or gene therapy to deliver cytokines or chemokines directly to the tumor site, thereby minimizing systemic side effects while concentrating immune-modulatory effects where they are most needed. Additionally, accumulating evidence supports combin ing cytokine or chemokine modulation with other immunother apies, particularly immune checkpoint inhibitors, to enhance therapeutic efficacy and overcome resistance mechanisms, as detailed in Table 1.
Table 1: List of cytokines and chemokines involved in cancer immunotherapy.
| Cytokine/Chemokine | Main Function in TME | Target Cell Types | Therapeutic Strategy |
|---|---|---|---|
| IFN-γ | Promotes anti-tumor immunity, enhances MHC expression | T cells, NK cells, tumor cells | Recombinant protein, gene therapy (Liao et al. 2019) |
| IL-2 | Activates T cells and NK cells | T cells, NK cells | Recombinant protein |
| IL-10 | Suppresses anti-tumor immunity, promotes Treg function | Tregs, macrophages | Monoclonal antibody (neutralizing) |
| TGF-β | Inhibits T cell activation, promotes MDSC differentiation | T cells, fibroblasts, MDSCs | Monoclonal antibody, small molecule inhibitor (Arce-Sillas et al. 2016) |
| CXCL12 | Promotes immunosuppressive cells, promotes tumor growth | Stromal cells, tumor cells | Receptor antagonist (e.g., CXCR4 inhibitor) (Li et al. 2020) |
| CCL2 | Recruit’s macrophages and MDSCs | Macrophages, MDSCs | Monoclonal antibody, small molecule inhibitor (Chang et al. 2016) |
Immune reprogramming in neurodegeneration
Neuroinflammation and immune system changes in neurodegenerative diseases
Neurodegenerative diseases, once viewed solely as disor ders of neuronal loss, are now recognized as having a substan tial inflammatory component. Neuroinflammation, defined as the inflammatory response within the Central Nervous System (CNS), represents a pathological hallmark of various disorders including Alzheimer’s Disease (AD), Parkinson’s Disease (PD), and Multiple Sclerosis (MS). This inflammatory state does not merely coincide with neuronal death but actively contributes to disease progression [62].
Microglia, the resident macrophages of the CNS, serve as the primary immune cells of the brain. In healthy brain tissue, microglia maintain a resting or homeostatic state, continuously surveying their microenvironment for pathogens or tissue dam age [63]. However, in neurodegenerative diseases, microglia undergo chronic activation, acquiring a reactive phenotype. This activation is triggered by various stimuli, including misfold ed protein aggregates such as amyloid-beta in AD and alpha synuclein in PD, as well as cellular injury [92].
Activated microglia can adopt a spectrum of functional states, ranging from a beneficial, phagocytic phenotype (M2 like) that clears debris and promotes tissue repair, to a detri mental, pro-inflammatory phenotype (M1-like) that secretes neurotoxic molecules and exacerbates neuronal injury. Astro cytes, another major glial cell population, also contribute to this neuroinflammatory milieu through release of pro-inflammatory mediators and disruption of synaptic function [103].
The role of cytokines and chemokines in neurodegeneration
The transition from a homeostatic to an inflammatory state in the CNS is orchestrated by a complex network of cytokines and chemokines. In neurodegenerative diseases, pro-inflamma tory cytokines including TNF-alpha, IL-1-beta, and IL-6 are con sistently elevated in patient brain tissue [1]. These molecules promote the pro-inflammatory phenotype of microglia and as trocytes, establishing a self-perpetuating cycle of inflammation and cellular damage. In AD, for instance, amyloid-beta plaques stimulate microglia to release these cytokines, which in turn ex acerbate plaque formation and neurotoxicity [64].
Chemokines play an equally critical role by directing the mi gration of both resident and peripheral immune cells. CCL2, for example, is elevated in AD and recruits peripheral monocytes into the CNS, potentially amplifying the inflammatory response [89]. Similarly, the CXCL12/CXCR4 chemokine axis has been im plicated in T cell infiltration into the CNS in MS, contributing to demyelination. The intricate interactions among these signaling molecules determine both the magnitude of inflammation and the specific immune cell populations that infiltrate the brain, ultimately influencing disease outcomes [2].
Therapeutic potential of cytokine and chemokine modulation in neurodegenerative disorders
Targeting cytokine and chemokine signaling pathways rep resents a promising therapeutic avenue for neurodegenerative diseases. Strategies aim either to inhibit detrimental pro-in flammatory responses or to enhance beneficial pro-repair re sponses. One approach involves neutralizing specific cytokines using anti-inflammatory agents [96]. For example, blocking TNF alpha or IL-1-beta has been investigated as a means to dampen the inflammatory cascade. However, the interconnected nature of the cytokine network suggests that broad anti-inflammatory strategies may not always prove effective and could potentially produce unwanted side effects [3,4].
An alternative strategy focuses on modulating chemokine re ceptors to prevent infiltration of pathogenic immune cells into the CNS [4]. Small molecule inhibitors of chemokine receptors have been designed to block immune cell migration. Addition ally, anti-inflammatory and neuroprotective cytokines, such as IL-4 and IL-13, are being explored for their potential to promote the beneficial M2-like microglial phenotype and enhance neu ronal survival. While still in early developmental stages, these approaches hold promise for establishing a new generation of therapies capable of directly modulating the neuroinflamma tory environment [6].
Mechanisms of cytokine and chemokine modulation
Cytokine signaling pathways in immune reprogramming
Cytokines exert their diverse biological effects by binding to specific cell surface receptors, initiating intracellular signal ing cascades that ultimately alter gene expression and cellular function. The Janus kinase/signal transducers and activators of transcription (JAK/STAT) pathway represents one of the most prominent cytokine signaling mechanisms [6]. Upon cytokine binding, receptor-associated JAK proteins become activated through phosphorylation, subsequently phosphorylating STAT proteins. These activated STAT proteins then dimerize, translo cate to the nucleus, and regulate transcription of target genes involved in immune cell differentiation, proliferation, and acti vation.
Alternative cytokine signaling pathways include the Nuclear Factor Kappa B (NF-κB) pathway and Mitogen-Activated Pro tein Kinase (MAPK) cascades [7]. The NF-κB pathway, activated by cytokines such as TNF-alpha and IL-1, plays a critical role in regulating inflammatory responses and cell survival. MAPK pathways, including ERK, JNK, and p38 MAPK, mediate diverse cellular responses including proliferation, differentiation, and apoptosis. The specific signaling pathway activated depends on both the cytokine involved and the cellular context, contribut ing to the remarkable versatility of cytokine-mediated immune regulation.
Understanding these signaling mechanisms is essential for developing targeted therapeutic interventions. For instance, JAK inhibitors have emerged as effective treatments for vari ous autoimmune and inflammatory conditions by blocking the downstream effects of multiple pro-inflammatory cytokines si multaneously [10]. Similarly, inhibitors targeting NF-κB or MAPK pathways are being investigated as potential therapies for both cancer and neurodegenerative diseases, where aberrant acti vation of these pathways contributes to disease pathogenesis [25].
Chemokine receptors and immune cell trafficking
Chemokines primarily signal through G Protein-Coupled Re ceptors (GPCRs), a large family of transmembrane receptors. Chemokine binding to its cognate receptor triggers activation of heterotrimeric G proteins, initiating downstream signaling cascades that predominantly affect cellular cytoskeletal dynam ics [9]. This leads to rapid reorganization of actin filaments and formation of a leading edge, enabling cells to migrate along chemokine concentration gradients [66]. This process, termed chemotaxis, plays a fundamental role in directing immune cells (including T cells, B cells, and neutrophils) to sites of inflamma tion, infection, or tumor formation, as illustrated in Figure 2.
Chemokine-receptor interactions exhibit remarkable speci ficity, ensuring that immune cell trafficking is highly regulated and directionally controlled [11]. However, this system also dis plays considerable redundancy, with multiple chemokines ca pable of binding the same receptor and individual chemokines binding multiple receptors. This complexity provides robustness to immune cell recruitment but also presents challenges for therapeutic targeting
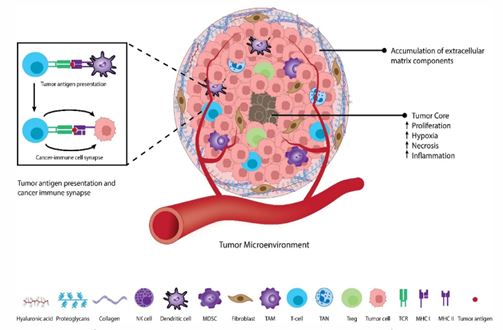
Table 2: Key cytokine and chemokine receptors and their roles.
| Receptor | Ligands | Associated Pathway | Key Role |
|---|---|---|---|
| IFNAR | IFN-α, IFN-β | JAK/STAT | Anti-viral and anti-proliferative response (Umansky et al. 2016) |
| TNFR | TNF-α, TNF-β | NF-κB, MAPK | Inflammation, cell death, cell survival (Siegel et al. 2022) |
| IL-2R | IL-2, IL-15 | JAK/STAT | T cell proliferation and differentiation (Galdiero et al. 2013) |
| CXCR4 | CXCL12 | GPCR | Immune cell trafficking, tumor metastasis (Fridlender & Albelda 2012) |
| CCR2 | CCL2 | GPCR | Macrophage and monocyte recruitment (Blattner et al. 2018) |
| IL-10R | IL-10 | JAK/STAT | Immunosuppression (Li et al. 2019) |
Notably, while IL-10 typically suppresses immune respons es, emerging evidence indicates it can paradoxically enhance tumor-resident CD8+ T cell activation in specific contexts [93].
The interplay between cytokine and chemokine signaling pathways is extensive and bidirectional [12]. Pro-inflammatory cytokines, including TNF-alpha and IL-1, can upregulate expression of various chemokine receptors and chemokines on both endothelial and immune cells, thereby establishing positive feedback loops that amplify immune cell recruitment to inflamed sites (Bellone et al. 2013). Similarly, certain cytokines can activate the JAK/STAT pathway, leading to transcriptional upregulation of chemokine receptors and enhancing immune cell responsiveness to chemokine gradients, as detailed in Table 2.
Cytokine and chemokine therapy in cancer treatment
Cytokine therapy for tumor immunotherapy
Cytokines were among the first therapeutic agents em ployed in cancer immunotherapy, with the goal of enhancing host anti-tumor immune responses. Recombinant cytokines such as Interferon-Alpha (IFN-alpha) and Interleukin-2 (IL-2) have received regulatory approval for treating specific malig nancies. IFN-alpha, one of the earliest immunotherapies, was initially approved for hairy cell leukemia and melanoma [13]. This cytokine exerts multiple anti-tumor effects, including direct inhibition of tumor cell proliferation, enhancement of antigen presentation through upregulation of MHC class I molecules, and augmentation of Natural Killer (NK) cell and T cell activity [14].
IL-2, approved for treatment of metastatic renal cell carci noma and metastatic melanoma, promotes proliferation and activation of T cells, particularly Cytotoxic T Lymphocytes (CTLs) and NK cells, which are essential for tumor cell destruction. These cells play critical roles in tumor immunosurveillance and elimination.
However, these first-generation cytokine therapies have been hampered by severe systemic toxicity. High-dose IL-2 can induce capillary leak syndrome, hypotension, and multi-organ failure due to systemic inflammatory response syndrome [95]. Similarly, IFN-alpha therapy is associated with flu-like symp toms, profound fatigue, and neurotoxicity. These adverse ef fects, coupled with the short half-lives of recombinant proteins, have prompted a shift toward more targeted and less toxic ther apeutic approaches [14].
Chemokine-based approaches to enhance anti-tumor immunity
As master regulators of immune cell trafficking, chemokines offer unique opportunities to reshape the TME for therapeutic benefit. Chemokine-based therapies aim either to recruit anti tumor immune cells to the tumor site or to prevent recruitment of pro-tumorigenic cells. One strategy involves direct delivery of chemokines to the tumor microenvironment [16]. For exam ple, gene therapy approaches have been explored to introduce genes encoding chemokines such as CXCL9 and CXCL10 into the TME. These chemokines attract CXCR3-expressing T cells and NK cells, which are highly effective at eliminating tumor cells.
This localized delivery strategy aims to create a chemokine gradient that recruits substantial numbers of anti-tumor im mune cells to the tumor, effectively converting an immunologi cally “cold” tumor into an inflamed, “hot” tumor. Conversely, an alternative approach employs chemokine receptor antagonists to block recruitment of immunosuppressive cells. For instance, inhibitors targeting CCR4, CCR5, and CCR6 have been investi gated to prevent regulatory T cell (Treg) migration to the TME, thereby reducing T cell suppression and enhancing anti-tumor immunity [17].
Clinical trials and successes in cytokine and chemokine therapy
Despite early setbacks, renewed interest in cytokine and chemokine therapies has emerged, particularly when used in combination with other treatment modalities. One promising strategy involves combining these agents with Immune Check point Inhibitors (ICIs) [18,19]. The rationale is that cytokine pre treatment can increase Tumor-Infiltrating Lymphocytes (TILs), which can then be activated by ICIs that release inhibitory sig nals. For example, IL-2 has demonstrated improved response rates when combined with ICIs in certain malignancies.
Next-generation cytokine-based therapies currently in clini cal trials include engineered cytokines with extended half-lives, fusion proteins combining cytokines with monoclonal antibod ies for tumor-targeted delivery, and cytokines delivered via oncolytic viruses [20]. A notable example is talimogene laher parepvec (T-VEC), an oncolytic virus expressing GM-CSF, which has been approved for melanoma treatment and demonstrates the clinical potential of this approach. Similarly, chemokine re ceptor antagonists are undergoing clinical evaluation for vari ous solid tumors, as summarized in Table 3 [21].
Table 3: Clinical trials using cytokine and chemokine therapy in cancer.
| Therapeutic agent | Mechanism of action | Clinical trial phase | Indication | Key findings/Status |
|---|---|---|---|---|
| Recombinant IL-2 (Aldesleukin) | Activates T cells and NK cells | Approved | Metastatic melanoma, renal cell carcinoma | Approved but high toxicity limits use (Sever et al. 2019) |
| Recombinant IFN-α | Enhances anti-tumor immunity, inhibits proliferation | Approved | Hairy cell leukemia, melanoma | Approved but associated with severe side effects (Raman et al. 2007) |
| T-VEC (Talimogene laherparepvec) | Oncolytic virus expressing GM-CSF | Approved | Melanoma | Improves durable response rates, especially in combination (De Sanctis et al. 2016) |
| Bempegaldesleukin (NKTR-214) | "Prodrug" of IL-2 that selectively activates T cells | Phase 3 | Melanoma, renal cell carcinoma | Designed to reduce systemic toxicity, but recent trials have shown mixed results (Gonzalez et al. 2018) |
| Anti-CCL2 monoclonal antibody | Blocks recruitment of macrophages to TME | Phase 2 | Various solid tumors | In trials to reduce immunosuppressive cell infiltration (Kartikasari et al. 2021) |
| CXCR4 antagonist (e.g., AMD3100) | Blocks tumor cell proliferation and metastasis | Phase 1/2 | Hematological and solid tumors | In trials to enhance chemo- and radiotherapy efficacy (Ozga et al. 2021) |
Cytokine and chemokine modulation in neurodegenerative diseases
Targeting inflammation and immune reprogramming in Alzheimer’s disease
The neuroinflammatory hypothesis of Alzheimer’s Disease (AD) posits that chronic inflammation represents a major driv er of disease progression, alongside the classical hallmarks of amyloid-beta plaques and neurofibrillary tangles [5]. Microglia and astrocytes primarily mediate this inflammatory process in AD through production of various pro-inflammatory cytokines, including TNF-alpha, IL-1-beta, and IL-6. Therapeutic strategies targeting AD aim either to reduce this chronic inflammatory state or to shift the microglial phenotype toward a more neuro protective profile [22].
One approach involves neutralizing pro-inflammatory cyto kines. TNF-alpha inhibitors, already approved for treating in flammatory conditions such as rheumatoid arthritis, have been evaluated in clinical trials for AD [23]. While early animal stud ies showed promise in reducing plaque burden and improving cognitive function, clinical trial results in human subjects have been inconsistent, with some studies showing minimal or no cognitive benefit. These challenges stem partly from the limited ability of these large molecules to cross the Blood-Brain Barrier (BBB) and the complex, context-dependent role that TNF-alpha plays in the CNS [24].
An alternative strategy leverages anti-inflammatory and neu rotrophic cytokines. IL-4, for instance, induces an anti-inflam matory, pro-phagocytic M2 phenotype in microglia. Preclinical studies have demonstrated that CNS delivery of IL-4 reduces amy loid-beta load and improves synaptic function. Similarly, IL-13 and Granulocyte Colony-Stimulating Factor (G-CSF) are being investi gated for their neuroprotective and neurogenic properties [25].
Cytokine and chemokine approaches in Parkinson’s disease and multiple sclerosis
Parkinson’s Disease (PD) involves progressive loss of dopa minergic neurons mediated by neuroinflammation, wherein activated microglia release pro-inflammatory cytokines and chemokines. Therapies aimed at modulating this inflammatory response are under active investigation [26]. For example, in hibition of the GM-CSF receptor has been explored as a strat egy to reduce microglial activation. Additionally, considerable research in PD has focused on the neurotrophic cytokine Glial Cell Line-Derived Neurotrophic Factor (GDNF) [101]. Although GDNF has demonstrated efficacy in preclinical models, its clini cal translation has been hindered by challenges in delivering this large protein to deep brain structures [20,27].
Multiple Sclerosis (MS), an autoimmune disease of the CNS characterized by T cell-mediated demyelination and neurode generation, has a more established history of cytokine-based therapies. Interferon-Beta (IFN-beta) has been a cornerstone MS treatment for decades [29]. Its mechanism of action is be lieved to be multifaceted, including suppression of pro-inflam matory cytokines, enhancement of anti-inflammatory cyto kines, and inhibition of T cell proliferation and migration across the BBB. The chemokine receptor CCR6 has also emerged as a therapeutic target, as its inhibition could prevent pathogenic T Helper 17 (Th17) cell migration into the CNS, a process impli cated in MS pathogenesis [30].
Challenges in therapeutic neurodegenerative diseases
Although cytokines and chemokines have been clearly im plicated in neurodegenerative disease pathogenesis, translat ing these findings into effective therapeutics has proven chal lenging. The most significant obstacle is the Blood-Brain Barrier (BBB), a highly selective semipermeable barrier that protects the brain from pathogens and potentially harmful blood-borne molecules [31]. While essential for brain homeostasis, the BBB severely restricts passage of large therapeutic molecules such as recombinant cytokines and monoclonal antibodies into the CNS. This limitation has compromised the efficacy of many promising agents in clinical trials for neurodegenerative diseas es, as outlined in Table 4.
Table 4: Potential cytokine and chemokine targets in neurodegenerative diseases.
| Therapeutic target | Mechanism of action | Disease | Clinical Trial Status | References |
|---|---|---|---|---|
| TNF-α | Neutralizes pro-inflammatory cytokine | Alzheimer's, Parkinson's | Mixed results in clinical trials due to BBB penetration and dual role | Altorki et al. 2019 |
| IL-1β | Neutralizes pro-inflammatory cytokine | Alzheimer's, Parkinson's | Preclinical and early-phase clinical trials | Wang et al. 2017 |
| GDNF | Promotes survival and function of dopaminergic neurons | Parkinson's | Clinical trials show limited success due to delivery challenges | Thunissen et al. 2014 |
| IFN-β | Modulates immune response, anti-inflammatory effects | Multiple Sclerosis | Approved for clinical use, a cornerstone therapy | Jacute et al. 2018 |
| CCR2/5 antagonist | Blocks recruitment of immune cells to CNS | Alzheimer's, MS | Preclinical and early-phase clinical trials | Liu et al. 2019 |
| IL-4/IL-13 | Promotes anti-inflammatory M2 microglial phenotype | Alzheimer's | Preclinical studies show promise | Siegel et al. 2019 |
To circumvent this barrier, researchers are exploring alterna tive delivery methods, including intracerebroventricular injec tions, engineered nanocarriers, and cell-based therapies. Addi tional strategies involve bispecific antibodies that bind both a BBB receptor and the therapeutic target, effectively hitchhiking across the barrier. These next-generation delivery systems are fundamental to realizing the full therapeutic potential of cyto kine and chemokine modulation in neurodegeneration [32].
Another major challenge lies in the complex, context-depen dent nature of cytokines and chemokines. The same molecule may exert beneficial or detrimental effects depending on tim ing, location, and concentration. For instance, an initial pro-in flammatory response may be beneficial for clearing misfolded proteins, whereas sustained inflammation causes irreversible neuronal damage. Broad suppression of inflammation, while seemingly logical, may disrupt protective immune functions such as pathogen clearance or debris removal, potentially in creasing vulnerability to other insults. Therefore, therapeutic modulation in neurodegeneration requires a nuanced approach that targets specific inflammatory pathways without causing global immune dysregulation [33,34].
Challenges and limitations of cytokine and chemokine therapy
Systemic toxicity and side effects in cancer treatment
While cytokine and chemokine modulation hold consider able therapeutic promise, significant obstacles have hindered clinical implementation, particularly regarding systemic toxicity and adverse effects. First-generation cytokine therapies, such as high-dose Interleukin-2 (IL-2) and Interferon-Alpha (IFN-alpha), exemplify these limitations [35]. Administration of these potent immune activators at therapeutic doses often triggers severe systemic inflammatory responses, resulting in narrow thera peutic windows.
Capillary leak syndrome, a life-threatening condition charac terized by fluid extravasation, severe hypotension, and multi organ dysfunction, commonly occurs in patients receiving high-dose IL-2. Similarly, IFN-alpha therapy is associated with debilitating flu-like symptoms, profound fatigue, and neurotox icity, frequently necessitating dose reduction or treatment dis continuation [36].
The primary contributor to this systemic toxicity is immune cell activation throughout the body rather than selective activa tion at the tumor site [98]. Although more recent approaches, including fusion proteins and oncolytic viruses, aim to achieve localized delivery, the risks of off-target effects and systemic inflammatory responses remain substantial concerns. This un derscores the critical need for advanced delivery systems and agents capable of selectively targeting pathological cell popula tions while sparing healthy tissues [37,38].
Context-dependent functions and immune system dysregulation in neurodegeneration
In neurodegenerative diseases, the challenges of cytokine and chemokine therapy are compounded by the delicate im mune balance within the Central Nervous System (CNS). Indi vidual molecules may exhibit dual or even multiple functions, with effects varying based on cellular context, disease stage, and local microenvironment [39]. A molecule may act as neu roprotective under certain conditions while proving neurotoxic in others. For instance, an initial pro-inflammatory response may prove beneficial for clearing misfolded protein aggregates, whereas chronic inflammation inevitably leads to irreversible neuronal damage [40].
Broad anti-inflammatory interventions, though seemingly rational, carry significant risks. Complete inhibition of major inflammatory cytokines such as TNF-alpha may compromise essential immune functions, including pathogen clearance and cellular debris removal, potentially increasing brain vulner ability to additional insults. Moreover, systemic immunosup pression could increase infection risk and impair tissue repair mechanisms. Consequently, therapeutic modulation in neuro degeneration demands precise, carefully calibrated approaches that target specific pathological inflammatory pathways while preserving physiological immune system functions [41].
SOvercoming delivery challenges in cytokine and chemokine therapy
The Blood-Brain Barrier (BBB) represents one of the most formidable obstacles for therapeutic agents targeting the CNS. This highly selective semipermeable barrier protects the brain from bloodborne pathogens and potentially harmful molecules [42,48,55]. However, this protective function simultaneously prevents large therapeutic molecules, including recombinant proteins and monoclonal antibodies, from reaching the CNS in therapeutically relevant concentrations. This limitation has severely restricted the clinical efficacy of numerous promising candidates in neurodegenerative disease trials [54].
To overcome this barrier, researchers are investigating in novative delivery strategies. These include nanocarrier systems engineered to transport therapeutic molecules across the BBB, as well as gene therapy and cell-based delivery approaches wherein genetically modified cells produce and secrete thera peutic cytokines or chemokines directly within the CNS [38]. Bispecific antibodies, designed to bind both BBB receptors and therapeutic targets, represent another promising strategy for facilitating transcytosis across the barrier. Additionally, focused ultrasound techniques can transiently disrupt the BBB, permit ting enhanced drug penetration. These next-generation deliv ery platforms are essential for unlocking the full therapeutic potential of cytokine and chemokine modulation in neurode generative diseases, and their continued development repre sents a critical research priority [53].
Conclusion
This comprehensive review has illuminated the pivotal and multifaceted roles of cytokines and chemokines in both cancer and neurodegenerative diseases. These signaling molecules function not as passive mediators but as active orchestrators of immune reprogramming that fundamentally drives disease progression. In cancer, malignant cells co-opt these pathways to establish an immunosuppressive microenvironment, recruiting pro-tumorigenic cells while simultaneously suppressing anti-tu mor immunity. Conversely, in neurodegeneration, dysregulated cytokine and chemokine signaling perpetuates chronic neuroin flammation, contributing to progressive neuronal damage and clinical deterioration.
We have examined diverse therapeutic approaches designed to modulate these pathways for clinical benefit, ranging from conventional recombinant cytokines to more sophisticated tar geted therapies including monoclonal antibodies and small mol ecule inhibitors. While significant progress has been achieved, particularly in cancer immunotherapy, substantial challenges remain. Systemic toxicity continues to limit the therapeutic win dow of many cytokine-based therapies, while the blood-brain barrier poses a formidable obstacle for CNS-targeted interven tions. Moreover, the context-dependent and often paradoxical functions of these molecules necessitate careful consideration of timing, dose, and delivery methods.
The future success of cytokine and chemokine-based ther apies depends critically on addressing these key challenges. Priority research areas should include development of highly specific agents capable of distinguishing pathological from physiological signaling. Personalized medicine approaches, uti lizing patient-specific biomarkers to tailor therapy and predict treatment response, will prove increasingly important. We an ticipate continued advancement in sophisticated delivery sys tems, including engineered nanocarriers and cell-based thera pies, to circumvent the blood-brain barrier and other delivery constraints.
Combination therapies, integrating cytokine and chemo kine modulation with other immunotherapies such as immune checkpoint inhibitors or CAR-T cell therapy, represent particu larly promising strategies with potential for synergistic effects. Additionally, temporal modulation strategies that account fordisease stage and dynamic changes in the immune microenvi ronment may enhance therapeutic efficacy while minimizing adverse effects. Ultimately, success will require deeper under standing of the spatiotemporal complexity and context-specific nature of immune signaling networks.
Final remarks
The fields of cancer immunotherapy and neurodegenera tion therapeutics are advancing at an unprecedented pace. While the path toward safe and effective treatments remains challenging, the potential to achieve therapeutic immune re programming through cytokine and chemokine modulation is increasingly tangible. As ongoing research continues to unravel the complexities of these signaling networks, we are approach ing a new era of precision medicine capable of restoring appro priate immune balance to combat these devastating diseases. The integration of cutting-edge technologies, including artificial intelligence for biomarker discovery, advanced biomaterials for targeted delivery, and sophisticated imaging techniques for real-time monitoring, promises to accelerate progress toward this goal. With continued interdisciplinary collaboration and sustained research investment, cytokine and chemokine-based therapies hold substantial promise for transforming clinical out comes in both cancer and neurodegenerative diseases.
References
- Marks ZR, Campbell NK, Mangan NE, Vandenberg CJ, Gearing LJ, Matthews AY, et al. Interferon-ε is a tumour suppressor and restricts ovarian cancer. Nature. 2023; 620: 1063–1070.
- Jacquelot N, Yamazaki T, Roberti MP, Duong CP, Andrews MC, Verlingue L, et al. Sustained type I interferon signaling as a mechanism of resistance to PD-1 blockade. Cell Res. 2019; 29: 846–861.
- Nan J, Wang Y, Yang J, Stark GR. IRF9 and unphosphorylated STAT2 cooperate with NF-κB to drive IL6 expression. Proc Natl Acad Sci U S A. 2018; 115: 3906–3911.
- Nakamura T, Sato T, Endo R, Sasaki S, Takahashi N, Sato Y, et al. STING agonist loaded lipid nanoparticles overcome anti-PD-1 resistance in melanoma lung metastasis via NK cell activation. J Immunother Cancer. 2021; 9: e002852.
- Ding L, Wang Q, Martincuks A, Kearns MJ, Jiang T, Lin Z, et al. STING agonism overcomes STAT3-mediated immunosuppression and adaptive resistance to PARP inhibition in ovarian cancer. J Immunother Cancer. 2023; 11: e005627.
- Lee SJ, Yang H, Kim WR, Lee YS, Lee WS, Kong SJ, et al. STING activation normalizes the intraperitoneal vascular-immune microenvironment and suppresses peritoneal carcinomatosis of colon cancer. J Immunother Cancer. 2021; 9: e002195.
- Ramanjulu JM, Pesiridis GS, Yang J, Concha N, Singhaus R, Zhang SY, et al. Design of amidobenzimidazole STING receptor agonists with systemic activity. Nature. 2018; 564: 439–443.
- Yi M, Niu M, Wu Y, Ge H, Jiao D, Zhu S, et al. Combination of oral STING agonist MSA-2 and anti-TGF-β/PD-L1 bispecific antibody YM101: a novel immune cocktail therapy for non-inflamed tumors. J Hematol Oncol. 2022; 15: 142.
- Yi M, Niu M, Zhang J, Li S, Zhu S, Yan Y, et al. Combine and conquer: manganese synergizing anti-TGF-β/PD-L1 bispecific antibody YM101 to overcome immunotherapy resistance in non-inflamed cancers. J Hematol Oncol. 2021; 14: 146.
- Pan BS, Perera SA, Piesvaux JA, Presland JP, Schroeder GK, Cumming JN, et al. An orally available non-nucleotide STING agonist with antitumor activity. Science. 2020; 369: eaba6098.
- Wu YT, Fang Y, Wei Q, Shi H, Tan H, Deng Y, et al. Tumor-targeted delivery of a STING agonist improves cancer immunotherapy. Proc Natl Acad Sci U S A. 2022; 119: e2214278119.
- Xu N, Palmer DC, Robeson AC, Shou P, Bommiasamy H, Laurie SJ, et al. STING agonist promotes CAR T cell trafficking and persistence in breast cancer. J Exp Med. 2020; 218: e20200844.
- Lu Q, Chen R, Du S, Chen C, Pan Y, Luan X, et al. Activation of the cGAS-STING pathway combined with CRISPR-Cas9 gene editing triggering long-term immunotherapy. Biomaterials. 2022; 291: 121871.
- Gorby C, Sotolongo Bellón J, Wilmes S, Warda W, Pohler E, Fyfe PK, et al. Engineered IL-10 variants elicit potent immunomodulatory effects at low ligand doses. Sci Signal. 2020; 13: eabc0653.
- Chard LS, Maniati E, Wang P, Zhang Z, Gao D, Wang J, et al. A vaccinia virus armed with interleukin-10 is a promising therapeutic agent for treatment of murine pancreatic cancer. Clin Cancer Res. 2015; 21: 405–416.
- Baganizi DR, Nyairo E, Duncan SA, Singh SR, Dennis VA. Interleukin-10 conjugation to carboxylated PVP-coated silver nanoparticles for improved stability and therapeutic efficacy. Nanomaterials. 2017; 7: 165.
- Lopes JE, Sun L, Flick HL, Murphy EA, Losey HC. Pharmacokinetics and pharmacodynamic effects of nemvaleukin alfa, a selective agonist of the intermediate-affinity IL-2 receptor, in cynomolgus monkeys. J Pharmacol Exp Ther. 2021; 379: 203–210.
- Pan Y, Hao Y, Han H, Chen T, Ding H, Labbe KE, et al. Nemvaleukin alfa, a novel engineered IL-2 fusion protein, drives antitumor immunity and inhibits tumor growth in small cell lung cancer. J Immunother Cancer. 2022; 10: e004913.
- Boyman O, Arenas-Ramirez N. Development of a novel class of interleukin-2 immunotherapies for metastatic cancer. Swiss Med Wkly. 2019; 149: w14697.
- Vaishampayan UN. Nemvaleukin alfa monotherapy and in combination with pembrolizumab in patients with advanced solid tumors: ARTISTRY-1. Clin Cancer Res. 2022.
- Tichet M, Wullschleger S, Chryplewicz A, Fournier N, Marcone R, Kauzlaric A, et al. Bispecific PD1-IL2v and anti-PD-L1 break tumor immunity resistance by enhancing stem-like tumor-reactive CD8+ T cells and reprogramming macrophages. Immunity. 2023; 56: 162–179.
- Piper M, Hoen M, Darragh LB, Knitz MW, Nguyen D, Gadwa J, et al. Simultaneous targeting of PD-1 and IL-2Rβγ with radiation therapy inhibits pancreatic cancer growth and metastasis. Cancer Cell. 2023; 41: 950–969.
- Ren Z, Zhang A, Sun Z, Liang Y, Ye J, Qiao J, et al. Selective delivery of low-affinity IL-2 to PD-1+ T cells rejuvenates antitumor immunity with reduced toxicity. J Clin Invest. 2022; 132.
- Niederlova V, Tsyklauri O, Kovar M, Stepanek O. IL-2-driven CD8+ T cell phenotypes: implications for immunotherapy. Trends Immunol. 2023; 44: 890–901.
- Codarri Deak L, Nicolini V, Hashimoto M, Karagianni M, Schwalie PC, Lauener L, et al. PD-1-cis IL-2R agonism yields better effectors from stem-like CD8+ T cells. Nature. 2022; 610: 161–172.
- Rosen DB, Kvarnhammar AM, Laufer B, Knappe T, Karlsson JJ, Hong E, et al. TransCon IL-2 β/γ: a novel long-acting prodrug with sustained release of an IL-2Rβ/γ-selective IL-2 variant. J Immunother Cancer. 2022; 10: e004991.
- Algazi AP, Twitty CG, Tsai KK, Le M, Pierce R, Browning E, et al. Phase II trial of IL-12 plasmid transfection and PD-1 blockade in immunologically quiescent melanoma. Clin Cancer Res. 2020; 26: 2827–2837.
- Hewitt SL, Bailey D, Zielinski J, Apte A, Musenge F, Karp R, et al. Intratumoral IL12 mRNA therapy promotes TH1 transformation of the tumor microenvironment. Clin Cancer Res. 2020; 26: 6284–6298.
- Zheng X, Wu Y, Bi J, Huang Y, Cheng Y, Li Y, et al. The use of supercytokines, immunocytokines, engager cytokines, and other synthetic cytokines in immunotherapy. Cell Mol Immunol. 2022; 19: 192–209.
- Ongaro T, Matasci M, Cazzamalli S, Gouyou B, De Luca R, Neri D, et al. A novel anti-cancer L19-interleukin-12 fusion protein with an optimized peptide linker. J Biotechnol. 2019; 291: 17–25.
- Strauss J, Heery CR, Kim JW, Jochems C, Donahue RN, Montgomery AS, et al. First-in-human phase I trial of a tumor-targeted cytokine (NHS-IL12) in metastatic solid tumors. Clin Cancer Res. 2019; 25: 99–109.
- Skrombolas D, Sullivan M, Frelinger JG. Development of an interleukin-12 fusion protein activated by matrix metalloproteinase 9. J Interferon Cytokine Res. 2019; 39: 233–245.
- Xue D, Moon B, Liao J, Guo J, Zou Z, Han Y, et al. A tumor-specific pro-IL-12 activates preexisting cytotoxic T cells to control established tumors. Sci Immunol. 2022; 7: eabi6899.
- Glassman CR, Mathiharan YK, Jude KM, Su L, Panova O, Lupardus PJ, et al. Structural basis for IL-12 and IL-23 receptor sharing. Cell. 2021; 184: 983–999.
- Makkouk A, Yang XC, Barca T, Lucas A, Turkoz M, Wong JT, et al. Off-the-shelf Vδ1 gamma delta T cells engineered with GPC-3-specific CAR and soluble IL-15. J Immunother Cancer. 2021; 9: e003441.
- Heczey A, Courtney AN, Montalbano A, Robinson S, Liu K, Li M, et al. Anti-GD2 CAR-NKT cells in relapsed or refractory neuroblastoma. Nat Med. 2020; 26: 1686–1690.
- Deng X, Shen Y, Yi M, Zhang C, Zhao B, Zhong G, et al. Combination of novel oncolytic herpesvirus with paclitaxel for breast cancer therapy. J Med Virol. 2023; 95: e28768.
- Guo J, Liang Y, Xue D, Shen J, Cai Y, Zhu J, et al. Tumor-conditional IL-15 pro-cytokine reactivates anti-tumor immunity. Cell Res. 2021; 31: 1190–1198.
- Zhang Y, Liao Q, Wen X, Fan J, Yuan T, Tong X, et al. Hijacking of the nervous system in cancer: mechanism and therapeutic targets. Mol Cancer. 2025; 24: 44.
- Baruch EN, Nagarajan P, Gleber-Netto FO, Rao X, Xie T, Akhter S, et al. Inflammation induced by tumor-associated nerves promotes resistance to anti-PD-1 therapy. Res Sq. 2023.
- Li W, Wu Z, Meng W, Zhang C, Cheng M, Chen Y, et al. Blockade of IL-6 inhibits tumor immune evasion and improves anti-PD-1 immunotherapy. Cytokine. 2022; 158: 155976.
- Soler MF, Abaurrea A, Azcoaga P, Araujo AM, Caffarel MM. New perspectives in cancer immunotherapy: targeting IL-6 cytokine family. J Immunother Cancer. 2023; 11: e007530.
- Amer H, Flanagan KL, Kampan NC, Itsiopoulos C, Scott CL, Kartikasari AE, et al. Interleukin-6 shapes the inflammatory tumor microenvironment in ovarian cancer. Cancers. 2025; 17: 1691.
- Schiller M, Ben-Shaanan TL, Rolls A. Neuronal regulation of immunity: why, how and where? Nat Rev Immunol. 2021; 21: 20–36.
- Salvador AF, de Lima KA, Kipnis J. Neuromodulation by the immune system: a focus on cytokines. Nat Rev Immunol. 2021; 21: 526–541.
- Baral P, Umans BD, Li L, Wallrapp A, Bist M, Kirschbaum T, et al. Nociceptor sensory neurons suppress neutrophil and γδ T cell responses in bacterial lung infections and lethal pneumonia. Nat Med. 2018; 24: 417–426.
- Pinho-Ribeiro FA, Verri WA, Chiu IM. Nociceptor sensory neuron–immune interactions in pain and inflammation. Trends Immunol. 2017; 38: 5–19.
- Klose CS, Mahlakõiv T, Moeller JB, Rankin LC, Flamar AL, Kabata H, et al. The neuropeptide neuromedin U stimulates innate lymphoid cells and type 2 inflammation. Nature. 2017; 549: 282–286.
- Wallrapp A, Riesenfeld SJ, Burkett PR, Abdulnour REE, Nyman J, Dionne D, et al. The neuropeptide NMU amplifies ILC2-driven allergic lung inflammation. Nature. 2017; 549: 351–356.
- Diaz-Salazar C, Bou-Puerto R, Mujal AM, Lau CM, von Hoesslin M, Zehn D, et al. Cell-intrinsic adrenergic signaling controls the adaptive NK cell response to viral infection. J Exp Med. 2020; 217: e20190549.
- Wieduwild E, Girard-Madoux MJ, Quatrini L, Laprie C, Chasson L, Rossignol R, et al. β2-adrenergic signals downregulate the innate immune response and reduce host resistance to viral infection. J Exp Med. 2020; 217: e20190554.
- Steinberg BE, Silverman HA, Robbiati S, Gunasekaran MK, Tsaava T, Battinelli E, et al. Cytokine-specific neurograms in the sensory vagus nerve. Bioelectron Med. 2016; 3: 7–17.
- Oetjen LK, Mack MR, Feng J, Whelan TM, Niu H, Guo CJ, et al. Sensory neurons co-opt classical immune signaling pathways to mediate chronic itch. Cell. 2017; 171: 217–228.
- Mamuladze T, Kipnis J. Type 2 immunity in the brain and brain borders. Cell Mol Immunol. 2023; 20: 1290–1299.
- Alves de Lima K, Rustenhoven J, Da Mesquita S, Wall M, Salvador AF, Smirnov I, et al. Meningeal γδ T cells regulate anxiety-like behavior via IL-17a signaling in neurons. Nat Immunol. 2020; 21: 1421–1429.
- Alves de Lima K, Rustenhoven J, Kipnis J. Meningeal immunity and its function in maintenance of the central nervous system in health and disease. Annu Rev Immunol. 2020; 38: 597–620.
- Tommasin S, Giannì C, De Giglio L, Pantano P. Neuroimaging techniques to assess inflammation in multiple sclerosis. Neuroscience. 2019; 403: 4–16.
- Shimizu S, Hirose D, Hatanaka H, Takenoshita N, Kaneko Y, Ogawa Y, et al. Role of neuroimaging as a biomarker for neurodegenerative diseases. Front Neurol. 2018; 9: 265.
- Nutma E, Willison H, Martino G, Amor S. Neuroimmunology—the past, present and future. Clin Exp Immunol. 2019; 197: 278–293.
- Zu Horste GM, Gross CC, Klotz L, Schwab N, Wiendl H. Next-generation neuroimmunology: new technologies to understand central nervous system autoimmunity. Trends Immunol. 2020; 41: 341–354.
- Wiendl H, Gross CC, Bauer J, Merkler D, Prat A, Liblau R. Fundamental mechanistic insights from rare but paradigmatic neuroimmunological diseases. Nat Rev Neurol. 2021; 17: 433–447.
- Eva L, Pleș H, Covache-Busuioc RA, Glavan LA, Bratu BG, Bordeianu A, et al. A comprehensive review on neuroimmunology: insights from multiple sclerosis to future therapeutic developments. Biomedicines. 2023; 11: 2489.
- Henry R, Deckert M, Guruviah V, Schmidt B. Review of neuromodulation techniques and technological limitations. IETE Tech Rev. 2016; 33: 368–377.
- Lewis PM, Thomson RH, Rosenfeld JV, Fitzgerald PB. Brain neuromodulation techniques: a review. Neuroscientist. 2016; 22: 406–421.
- Abdul-Rahman T, Ghosh S, Badar SM, Nazir A, Bamigbade GB, Aji N, et al. The paradoxical role of cytokines and chemokines at the tumor microenvironment. Eur J Med Res. 2024; 29: 124.
- Yi M, Li T, Niu M, Zhang H, Wu Y, Wu K, et al. Targeting cytokine and chemokine signaling pathways for cancer therapy. Signal Transduct Target Ther. 2024; 9: 176.
- Altorki NK, Markowitz GJ, Gao D, Port JL, Saxena A, Stiles B, et al. The lung microenvironment: an important regulator of tumour growth and metastasis. Nat Rev Cancer. 2019; 19: 9–31.
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019; 69: 7–34.
- Thunnissen E, Noguchi M, Aisner S, Beasley MB, Brambilla E, Chirieac LR, et al. Reproducibility of histopathological diagnosis in poorly differentiated NSCLC. J Thorac Oncol. 2014; 9: 1354–1362.
- Jackute J, Zemaitis M, Pranys D, Sitkauskiene B, Miliauskas S, Vaitkiene S, et al. Distribution of M1 and M2 macrophages in non-small cell lung cancer. BMC Immunol. 2018; 19: 3.
- Liu J, Li Y, Zhu X, Li Q, Liang X, Xie J, et al. Increased CX3CL1 mRNA expression as a prognostic factor in lung adenocarcinoma. Oncol Lett. 2019; 17: 4877–4890.
- Wang M, Zhao J, Zhang L, Wei F, Lian Y, Wu Y, et al. Role of tumor microenvironment in tumorigenesis. J Cancer. 2017; 8: 761.
- Sever T, Leblebici A, Koc A, Binicier C, Olgun A, Edizer DT, et al. New insight of tumor microenvironment in non-small cell lung cancer. J Basic Clin Health Sci. 2019; 3: 113–120.
- Liao Z, Tan ZW, Zhu P, Tan NS. Cancer-associated fibroblasts in tumor microenvironment. Cell Immunol. 2019; 343: 103729.
- Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer. Genes Dev. 2018; 32: 1267–1284.
- Kartikasari AE, Huertas CS, Mitchell A, Plebanski M. Tumor-induced inflammatory cytokines and emerging diagnostic devices. Front Oncol. 2021; 11: 692142.
- Raman D, Baugher PJ, Thu YM, Richmond A. Role of chemokines in tumor growth. Cancer Lett. 2007; 256: 137–165.
- Umansky V, Blattner C, Gebhardt C, Utikal J. The role of myeloid-derived suppressor cells in cancer progression. Vaccines. 2016; 4: 36.
- De Sanctis F, Solito S, Ugel S, Molon B, Bronte V, Marigo I. MDSCs in cancer. Biochim Biophys Acta Rev Cancer. 2016; 1865: 35–48.
- Ozga AJ, Chow MT, Luster AD. Chemokines and the immune response to cancer. Immunity. 2021; 54: 859–874.
- Li BH, Garstka MA, Li ZF. Chemokines and recruitment of myeloid-derived suppressor cells. Mol Immunol. 2020; 117: 201–215.
- Chang AL, Miska J, Wainwright DA, Dey M, Rivetta CV, Yu D, et al. CCL2 produced by glioma microenvironment recruits Tregs and MDSCs. Cancer Res. 2016; 76: 5671–5682.
- Blattner C, Fleming V, Weber R, Himmelhan B, Altevogt P, Gebhardt C, et al. CCR5+ myeloid-derived suppressor cells in melanoma. Cancer Res. 2018; 78: 157–167.
- Li A, Herbst RH, Canner D, Schenkel JM, Smith OC, Kim JY, et al. IL-33 signaling alters regulatory T cell diversity. Cell Rep. 2019; 29: 2998–3008.
- Arce-Sillas A, Álvarez-Luquín DD, Tamaya-Domínguez B, Gomez-Fuentes S, Trejo-García A, Melo-Salas M, et al. Regulatory T cells in immune regulation. J Immunol Res. 2016; 2016: 1720827.
- Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin. 2022; 72.
- Galdiero MR, Garlanda C, Jaillon S, Marone G, Mantovani A. Tumor-associated macrophages and neutrophils. J Cell Physiol. 2013; 228: 1404–1412.
- Fridlender ZG, Albelda SM. Tumor-associated neutrophils: friend or foe? Carcinogenesis. 2012; 33: 949–955.
- Uribe-Querol E, Rosales C. Neutrophils in cancer. J Immunol Res. 2015; 2015: 983698.
- Zenobia C, Hajishengallis G. Interleukin-17 in immunity and inflammation. Periodontol 2000. 2015; 69: 142–159.
- Mano Y, Shirabe K, Yamashita YI, Harimoto N, Tsujita E, Takeishi K, et al. Neutrophil-to-lymphocyte ratio predicts survival after hepatectomy. Ann Surg. 2013; 258: 301–305.
- Luangdilok S, Box C, Harrington K, Rhŷs-Evans P, Eccles S. MAPK and PI3K signaling regulate cytokine secretion. Eur J Cancer. 2011; 47: 520–529.
- Jin L, Han B, Siegel E, Cui Y, Giuliano A, Cui X. Breast cancer lung metastasis. Cancer Biol Ther. 2018; 19: 858–868.
- Lillard J, Singh R, Sharma P, Singh S. CXCL13 inhibition prevents bone metastasis in prostate cancer. J Immunol. 2010; 184: 133–138.
- Mantovani A, Barajon I, Garlanda C. IL-1 and IL-1 regulatory pathways in cancer. Immunol Rev. 2018; 281: 57–61.
- Cursons J, Souza-Fonseca-Guimaraes F, Foroutan M, Anderson A, Hollande F, Hediyeh-Zadeh S, et al. Gene signature predicting NK cell infiltration in melanoma. Cancer Immunol Res. 2019; 7: 1162–1174.
- Moreira A, Alari-Pahissa E, Munteis E, Vera A, Zabalza A, Llop M, et al. Adaptive features of natural killer cells in multiple sclerosis. Front Immunol. 2019; 10: 2403.
- Iraolagoitia XLR, Spallanzani RG, Torres NI, Araya RE, Ziblat A, Domaica CI, et al. NK cells restrain antitumor CD8+ T cell priming. J Immunol. 2016; 197: 953–961.
- Bellone M, Calcinotto A. Enhancing lymphocyte trafficking into tumors. Front Oncol. 2013; 3: 231.
- Isvoranu GG, Surcel M, Munteanu AN, Bratu OG, Ionita-Radu F, Neagu MT, et al. Therapeutic potential of interleukin-15 in cancer. Exp Ther Med. 2021; 22: 675.
- Chow MT, Ozga AJ, Servis RL, Frederick DT, Lo JA, Fisher DE, et al. CXCR3 chemokine system required for anti-PD-1 efficacy. Immunity. 2019; 50: 1498–1512.
- Tan HY, Yip LY. The tumor microenvironment as a target for chemoprevention. Cancer Prev Res. 2018; 11: 449–462.
- Holicek P, Rackova L, Sobotka O. Astrocyte-neuron interactions in amyotrophic lateral sclerosis. Biomed Pharmacother. 2024; 170: 116045.